Siderophores are iron chelating compounds secreted by microorganisms to overcome the limitations and toxicity of iron for the regulation, storage and acquisition of heme uptake, degradation, Reductive iron assimilation. Aspergillus fumigatus is a saprophytic, air borne fungal pathogen of humans. A crucial role played by this fungal siderophore in a murine model of asperigillosis of the biosynthetic pathway for the virulence. A. fumigatus secretes Fusarinines C (FsC), Triacetylfusarinine C (TAFC) siderophores together extracellularly for the uptake and mobilization and Ferricrocin (FC), Hydroxyferricrocin (HFC) intracellularly for the storage and distribution of iron. Hydroxylation of orthinine is the precursor of arginine is involved in biosynthesis of fungal siderophores. Metabolism, Sensing of iron, uptake and regulation factors in A. fumigatus is not only through the two central transcription factors connected to a negative loop, GATA (SreA) and bZip (HapX) but also through the srbA and ergosterol biosynthesis. Deactivation in either of genes confines growth during iron starvation and sufficiency. For Iron transporting systems in fungi adopts four different mechanisms for the chelation of fe III ions and make it available in ferrous form for the appropriate absorption and function in plants.
Keywords
A. fumigatus
Fungal siderophores
Iron
Orthinine
Ergosterol
Transcriptional factors
Biosynthesis.
Important Note
Key findings:
Siderophores, including Fusarinines C (FsC) and Triacetylfusarinine C (TAFC), play a crucial role in Aspergillus fumigatus's virulence by regulating iron acquisition and distribution. Orthinine hydroxylation is key in siderophore biosynthesis. Iron metabolism in A. fumigatus involves GATA (SreA) and bZip (HapX) transcription factors, srbA, and ergosterol biosynthesis, crucial for growth under varying iron conditions.
What is known and what is new?
Siderophores are known iron-chelating compounds secreted by microorganisms to regulate iron acquisition and storage. Aspergillus fumigatus, a fungal pathogen, produces siderophores like Fusarinines C (FsC) and Triacetylfusarinine C (TAFC) for iron uptake and storage, crucial for its virulence. This study adds to this knowledge by highlighting the significance of orthinine hydroxylation in siderophore biosynthesis in A. fumigatus. It also identifies key transcription factors (GATA/SreA, bZip/HapX), srbA, and ergosterol biosynthesis in regulating iron metabolism in A. fumigatus, crucial for growth under varying iron conditions.
What is the implication, and what should change now?
The implications of this study are significant for understanding how A. fumigatus regulates iron metabolism, which is crucial for its virulence. This knowledge could lead to the development of new strategies for combating A. fumigatus infections, such as targeting key iron metabolism pathways. Additionally, the study underscores the importance of considering the complex mechanisms involved in iron regulation in fungi, which could have broader implications for understanding fungal pathogenesis and developing antifungal therapies.
Introduction
Aspergillus fumigatus an omnipotent saprophytic fungus resides mainly in decaying organic matter or in soil and behaves as a recycler in nature. This belongs to a flexible metabolism under the environmental conditions which meets its nutritional requirements (Kwon-Chung et al., 2013) [1] and is isolated from human habitats (Hirsch et al.,2000) [2] and vegetable compost heaps (Beffa et al., 1998 andRyckeboer et al., 2003) [3,4].
Now-a-days, A. fumigatus is considered as a weak pathogen causing allergic diseases, observed among individuals like farmers lung exposed to aspergilloma or conidia (Dixon et al., 1992; Kwon Chung et al., 1992 [5,6] and Pennington., 1988) [7]. It causes many disorders effecting human organs like immune system, immune compromised hosts, lungs, liver, brain and central nervous system. These disorders may cause many diseases in humans such as asthma, sinusitis, ethimoid sinuses, subsequent cranial bone osteomylitis, intracranial extension, episoidic wheezing, pulmonary infiltrates, sputum and blood eosinophilia, ABA, asperigillosis, Leukemia, cystic fibrosis, and tuberculosis (Table 1).
Table 1: Diseases Caused By A. Fumigates Affecting Different Human Organs
Human organs
Type of disease caused by
A. fumigatus
References
Immune system
asthma and sinusitis
Zureik et al.,(2002) [8]
Immunocompromised
hosts
Subsequent cranial bone osteomyelitis, intracranial extension and ethmoid sinuses.
Grigoriv et al.,(1979) [9]; Rudwan et al.,(1976) [10]; Rothfield et al., (1972) [11]
Lungs
episodic wheezing, pulmonary infiltrates, sputum and blood eosinophilia, pyrexia, and sputum containing brown flecks or plugs
Rosenberg et al.,(1977) [12]
Lungs
Allergic bronchopulmonary aspergillosis (ABA)
Agarwal et al.,(2013) [13]
solid organ transplant
recipients and/ or bone marrow
Aspergillosis
Wald et al.,(1997) [14]
Lungs, liver or brain
chronic granulomatous, invasive aspergillosis
Segal et al.,(1998) [15]
Central nervous system
If pulmonary aspergillosis
spreads, the mortality rate reaches to 100 % from 65%
De Bock., (1994) [16]
Lungs (bronchopulmonary disorders)
Leukemia, cystic fibrosis, and tuberculosis
Marr and Denning., (2002) [17]
A. Fumigatus can grow in 80% of human serum media, this is essential and possess an efficient mechanism for acquiring of iron [18].
Pulmonary disease is represented by conidial germination in the spore inhalation and human lungs. During spore germination, three crucial steps are distinguished they are as follows:
(i). At environmental conditions, activation of resting spore
(ii). isotropic growth that is responsible in growth of the wall and uptake of water
(iii).The new mycelium originates from the germ tube formation, as this is involved in the polarized growth (d’ Enfert., 1997 and Momany., 2002) [19].
cell wall of A. fumigatus is composed of a fibrillar branched β1,3-glucan core bound to chitin, galactomannan and β1,3-1,4-glucan, embedded in an amorphous cement composed of α1,3-glucan, galactomannan and polygalactosamine [20]. Hyphal cell wall is crucial to host cell resistance and for penetration of solid nutrient substrates by A. fumigatus.Homologues of the yeast PIR proteins face scantiness by A. fumigatus, which are credibly bound to β1,3-glucans by an alkali-labile bond [20]. This genome consists of 26 type of clusters with non-ribosomal peptide synthase, polyketide synthase with and/or without dimethylallyl tryptophan synthase genes [21].
In animals, A. fumigatus causes aspergillosis in serum, neutropenic, urinary excretion, was observed in guinea pigs, mice and rabbit (Table 2).The detection methods for the
Infected fumigatus are in urine via immunoassays in guinea pigs, from fumigatus strain (90906) in mice, antigen in urine and serum by counter immune electrophoresis in rabbit.
Table 2: Disease Occurrence with Infected A. Fumigatus in Animals and Its Detection Methods
Animals
Presence of Disease infected with A. fumigatus
Detection
References
Guinea pigs
serum
in urine via immunoassays
Dufresne et al., (2012) [22]
Mice
neutropenic
A. fumigatus (strain 90906)
Stephens-Romero et al., (2005) [23]
Rabbit
urinary excretion, model of experimental systemic aspergillosis
antigen in urine and serum by counter-immuno-electrophoresis
Lehmann et al., (1978) [24]
A. Fumigatus is considered as air borne fungal pathogen of humans, (Tekaia et al., 2005) [25] and a role of genome sequence. Recently A. Fumigatus is considered as one of the best studied fungal filamentous species. This genome sequence permit molecular analysis and allows “genome mining” (Nierman et al., 2005) [26]. However, A. fumigatus lacks particular uptake systems for iron host sources as transferrin heme and/ or ferritin (Schrettl et al., 2004) [27].
2. Requirement Iron by A. Fumigatus:
For all prokaryotes nearly and to all eukaryotes, iron is an essential nutrient [28]. In Earth’s crust, it is the fourth most opulent transition element [29] that can present in two oxidation states, they are as: Fe (II) Fe (III). The general functions of metabolic and cellular process for the requirement of iron occurs at, electron transport chain, tricarboxylic acid cycle amino acid metabolism andoxidative phosphorylation in A. fumigatus. Iron helps in nucleic acid synthesis and biosynthesis of vitamins, porphyrins, toxins, cytochromes, pigments, aromatic compounds, siderophores, and antibiotics (Table 3).
At physiological pH level (7.35–7.40), the ferric form of iron Fe (III) is insoluble and ferrous form of iron Fe (II) is soluble [30]. The variable valence of the iron allows it to act as a major role in the oxidation reduction reactions [31]. Iron at excess conditions catalyzes the formation of ROS that can damage cellular components (Table 3), on the other hand hydrophobicity of microbial surface reduces, which changes the surface protein composition and leads to the formation of biofilm at low conditions. There are some mechanisms for balancing storage, consumption and acquisition of iron in order to overcome limitation and excess amounts of iron (Table 3).
Table 3: Requirement of Iron in A. fumigatus
Functions
Occurs
References
Metabolic processes
electron transport chain tricarboxylic acid cycle, photosynthesis and oxidative phosphorylation,
Messenger and Barclay (1983); Fardeau et al., (2011). [32,33]
formation of cell damaging reactive oxygen species that can damage cellular components
Halliwell et al., (1984).[35]
Iron at limiting conditions:
Decreases
The biofilm formation is limited due to the decreasing of surface hydrophobicity which is altered by surface protein composition.
Simões et al., (2007). [36]
To overcome limitation of iron and to avoid excess iron:
Mechanisms
to balance the strategies of storing, acquisition and iron consumption.
Schrettl and Hass., (2011). [37]
3. Siderophores Production by A. fumigatus:
Mono and/or di iron are assimilated into heme sulfur clusters, as this metal is an obligatory co-factor for biosynthesis of DNA, cellular process and sterols varieties [34]. The limitation of iron availability increases in atmospheric oxygen which is owed to its oxidation into the insoluble ferric hydroxides [38]. Microorganisms have developed few particular strategies like siderophore production, because of the limited availability of iron in the environment [39].
Siderophores (In Greek, sideros means “iron” and phores means “bearer”) are the metal chelating compounds that functions mainly to encapsulate the insoluble ferric iron from environment [40].
Classification of hydroxamate siderophores in A.Fumigatus:
A. Fumigatus produce hydroxamate group of siderophores. Siderophores of Biosynthesis in A. Fumigatus is byornithine hydroxylation (orn) catalysed by the ornithine monooxygenase Sid A. For siderophore biosynthesis 40-phosphopantetheinyl transferase PptA is crucial because lysine-biosynthetic a-aminoadipate reductase, NRPS along with polyketide synthetases involves activation through this enzyme [41,42].
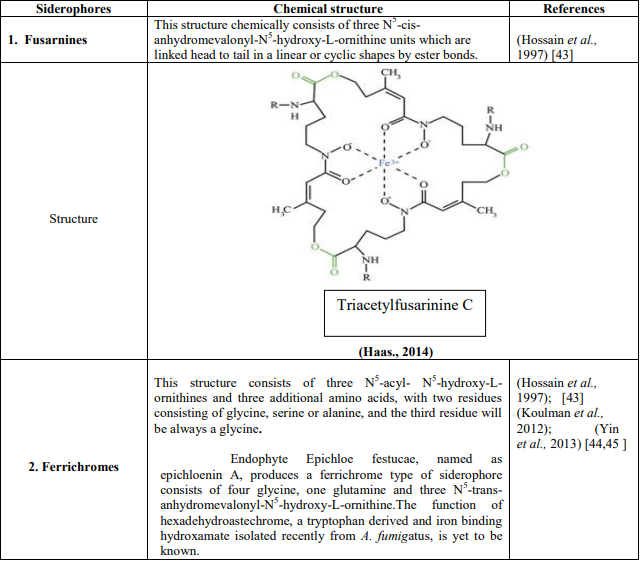
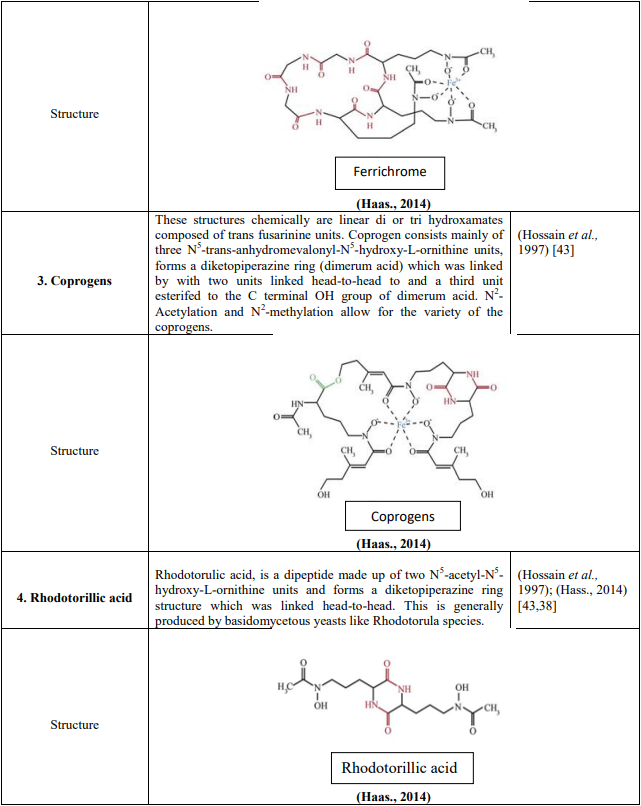
In A. Fumigatus, 4 types of siderophores are secreted (Table 5). Two in extracellular namely fusarinines C (Fs C) and triacetylfusarinineC (TAFC). Two in intracellular namely hyphal ferricrocin (FC) and hydroxyferricrocin (HFC)
In FsC R =H, all siderophores were represented in ferric forms, except rhodotorulic acid., and Fe (III), ester bonds and peptide bonds were displayed in blue, green and red colours respectively in Table 4.
Table 4: Chemical compounds and structures of four types of siderophores produced by A. fumigatus
4. Siderphore biosynthesis in A. fumigatus:
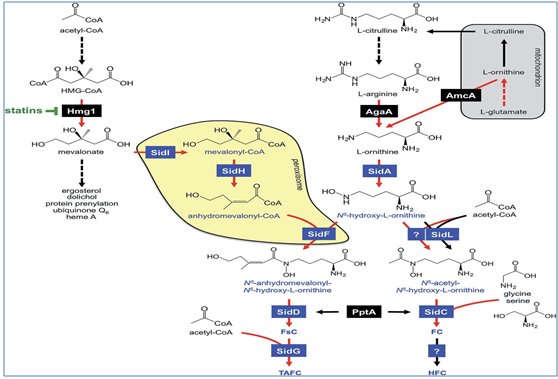
Ornithine monooxygenase SidA which catalyses the hydroxylation of ornithine is the prior catalytic step in all four types of siderophore biosynthesis. The splitting of different aceyl groups that transfers to N5-hydroxyornithine [41,42]. In mitochondrion, the first essential precursor ornithine is produced and all its biosynthetic pathway export through the transporter AmcA to the cytosolis were upregulated transcriptionally under iron deprivation in [46].
Conversion of arginine to ornithine by the arginase AgaA in response to iron starvation is converted in the cytosol, as ornithine is a precursor of arginine. SidF transfers, the transacylase anhydromevalonyl to N5-hydroxy-L-ornithine in the extracellular siderophores represented in (Fig 2) [47]. Two transacetylases like SidL and an unknown enzyme are required for the intracellular ferrichrome type of siderophores for their upregulation by starvation of iron [48].
Fusarinines and ferrichromes are formed primarily by Sid D and Sid C, two different non ribosomal peptide synthetases (NRPSs), an enzyme which is best recognized from secondary metabolism in A. fumigatus [47]. “GCN5-related N-acetyltransferases (GNAT)” proteins comes under SidF and SidL. lysine-biosynthesis involves in Polyketide synthetases similar to NRPSs, and the a-aminoadipate reductase, this depends on Sfp-type 40-phosphopantetheinyl transferase activation, entitled as PptA [42].
Similar to fusarinine, biosynthesis of ferrichrome A is also connected to biosynthesis of isoprenoide as methylglutaconyl-CoA which is derived from HMG-CoA through the enoyl-CoA-hydratase Fer4 dehydration [49]. As for TAFC biosynthesis mevalonate acts as a precursor and produces HMG-CoA reductase Hmg1. Additionally, the over expression of mevalonate increases production of TAFC. The inhibition of lovastin mediated Hmg 1 blocks biosynthesis of TAFC. At isoprenoid biosynthetic pathway, mevalonate acts as an intermediate with ergosterol as significant product, this pathway is helpful in acting against fungal diseases, as it increase resistance in amphotericin B and azoles. Whereas, iron deficiency reduces the level of cellular ergosterol due to the iron requirement for biosynthesis of ergosterol [50].
SidG is important for the TAFC biosynthesis. This deficiency of sidG, removes production of TAFC with increase in the production of FsC, and neither affects virulence nor growth [47] ∆sidI, sidH, sidF, sidD (Extracellular) and ∆sid C mutants (Intracellular) causes partial attenuation of virulence (Table 5).
With the only hydroxylation, HFC is derived from FC which is catalyzed by an unknown gene product shown in (Fig 2) (Schrettl et al., 2007) [47]. Both the extra and intra cellular siderophores are important for virulence, because the entire SB (ΔsidAmutant) elimination causes in supreme avirulence of invasive pulmonary aspergillosis in a murine model of A. fumigatus (Table 5) [51, 52].
Fig 2:Siderphore BiosynthesisPathway in A. fumigatus indicates in blue color and it is attached to the isoprenoide biosynthesis and also ornithine and/ or arginine metabolism. Enzymes involved are described briefly in the text box. During iron starvation, enzymatic steps upregulated transcriptionally and marked by red arrows. A broken arrow displays reactions involving enzymes more than one. Peroxisome and mitochondrion are shaded in grey and yellow, respectively [53].
Table 5: Siderophores produced by A. fumigatus
Extracellular siderophores
Intracellular siderophores
fusarinines C (FsC):
triacetylfusarinine C (TAFC):
hyphal ferricrocin (FC):
hydroxyferricrocin (HFC):
Chemical compound
Fs C is a cyclic tripeptide linked by ester bonds consists of three N5anhydromevalonyl N5 hydroxyornithine residues (Schrettl et al., 2007). [54]
TAFC is triple N2-acetylation of FsC catalyzed by SidG (Schrettl et al., 2007).
[54]
FC is a cyclic hexapeptide with the Gly Ser-Gly-(N5- acetyl-N5 hydroxyornithine)3 (Schrettl et al., 2007). [54]
HFC (∆sidC and ∆sidL, mutants) is hydroxylated FC. (Schrettl et al., 2007). [54]
catalyzation
FsC (extracellular) and FC (intracellular) are assembled and catalyzed by SidD and SidC i.e., two different non-ribosomal peptide synthetase (NRPS).
TAFC and HFC, both extracellular and intracellular siderophores are shaped by SidG-mediated N2-acetylation of FsC and hydroxylation of FC.
Genetic elimination
Under iron limitation, it decreases growth, resistance of oxidative stress, conidiation and few other iron acquisition systems are also responsible in reducing the lacking of siderophores.
Blocks sexual development and diminishes conidiation. The reduction in the supply of iron responsible for conidial iron shortage, by catalase A enzyme which impairs as iron dependent and consequently decreases conidial resistance to oxidative stress. Besides, during iron starvation, such type of conidia germination delays
Deficiency
(∆sidI, ∆sidH, ∆sidF or ∆sidD mutants) causes partial attenuation of virulence.
(∆sidC mutants) causes partial attenuation of virulence. For release of iron, TAFC and FsC are hydrolyzed to some extent by the esterase EstB (Kragl et al., 2007) [55]. In A.fumigatus, deficiency of ferricrocin decreases its germination and conidia iron content, oxidative stress resistance of hyphae (Eisendle et al., 2006 [56] and Schrettl et al., 2007)[54] and increases the labile iron pool (Eisendle et al., 2003 and Schrettl et al., 2007) [56,54]
NRPS links enzymes
SidD links three N5-cis anhydromevalonyl-
N5-hydroxy-L-ornithine residues by ester bonds, yields FsC (Schrettl et al., 2007) [54]
SidC links three N5-acetyl-N5-hydroxy-
L-ornithine, two L-glycine and one L-serine residue yields hexapeptide ferricrocin through peptide bonds (Schrettl et al., 2007) [54]
5. Iron homeostasis in A.fumigatus:
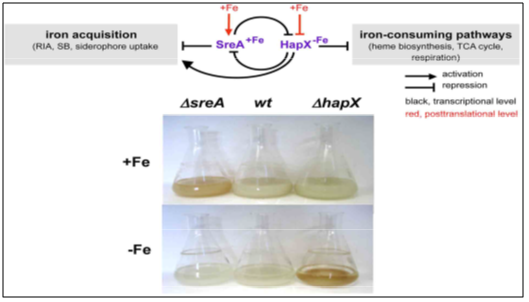
Iron homeostasis inculpate the two central transcription factors (TF) they are (i) GATA factor- SreA and (ii) bZip factor- HapX (Haas et al., 1999; Hortschansky et al., 2007; Schrettl et al., 2008, 2010 a) [57-60]. During iron sufficiency, SreA presume by bioinformatic analyses to identify ATCWGATAA sequences, in order to represses iron toxicity, high affinity iron uptake, including reductive iron assimilation and the siderophore system [59]. During iron deficiency, HapX decreases iron consuming pathways such as respiration, heme biosynthesis, tricarboxylic cycle and ribosome biogenesis to reserve iron shown in (Table 6).
Moreover, siderophores and ribotoxin AspF1 synthesis is activated by HapX and furtherly by coordination of siderophore biosynthesis with the supply of ornithine as its precursor in A. fumigatus. There is an interconnection loop between the two central transcriptional factors (SreA) and (HapX). During iron deficiency HapX repress sreA and while during iron sufficiency SreA represses hapX, where as both factors are regulated by iron blocking HapX function and activation of SreA function post translationally. Oninactivation of either SreA or HapX factors, the malignant effects are cramped growth during sufficiency and/ or deficiency with their mode of action and expression pattern is shown in (Fig 3) respectively.
A. fumigates consists of 24 genes which are having functional and identical characterization increases in iron that tends to cause respiration and tri carboxylic acid cycle as iron dependent [61]. Analysis of Proteome by A. fumigatus reveals that with increasing the contents of heme, cellular iron, zinc and copper, increases the protein production in tricarboxylic acid cycle, glycolysis and respiration in hypoxia [62].
SrbA regulates iron uptake in response to adaptation of hypoxia and in most eukaryotes it is sustained and activates siderophore and transcriptional factor of sterol regulatory element binding protein “(SREBP)” [61]. To maintain sterol homeostasis, cellular sterol exhaustion activates these TFs (Bien and Espenshade., et al., 2010); (Blatzer et al., 2011 a) [63, 64]. Deficiency of SrbA decreases the content of cellular ergosterol, resistance against azole drugs and hypoxic growth is arrested as well as virulence [61].
During iron starvation, SrbA transcriptional activation is independent of SreA and HapX factors. So, this states that A. fumigatus recognizes iron not only through theHapX and SreA but also through SrbA and sterol biosynthesis. Whereas deficiency of SreA leads to the excessive uptake of iron and upregulates both the enzymes transcriptionally [65]. The shortage in SreA or HapX both the factors leads to reddish hyphal pigmentation caused by metabolic deregulation. Only deficiency in HapX of bZip factor occurs A. fumigatus virulence of aspergillosis in murine models [59,60], which draws attention in the limitation stages of iron adaptation role (Table 6).
In order to maintaincell wall integrity, secondary metabolism, protection against reactive oxygen species and mitogen activated protein kinase (MAPK), iron starvation triggers nuclear localization and phosphorylation in A. fumigatus whereas MpkA plays a major role (Jain et al., 2011) [66] and this deficiency increases production of siderophore.
Fig 3: Regulation of iron homeostatis in A. fumigates (A) SreA and HapX mediated iron regulation. (B) A. fumigatus Phenotypes i.e., SreA (Δ sreA) and HapX (Δ hapX) deficient flask mutant strains in 24h/37˚C cultures. During iron starvation, in contrast to the wild type (wt), due to accumulation of protoporphyrin IX, Δ hapX mycelia are reddish colored and in iron sufficiency, Δ sreA mycelia are reddish colored due to accumulation of heme, iron, and FC (Schrettl et al., 2008, 2010a) [59,60].
Table 6: Regulatory functions of Central transcriptional factors involved in Iron Homeostasis of A. fumigates:
Regulatory functions of A. fumigates
Central Transcription Factors
1. GATA factor - SreA
2. bZip factor– HapX
At iron sufficiency
( Fe+)
To avoid iron toxicity, SreA represses high affinity uptake of iron, including siderophore system and Reductive Iron Assimilation.
Ribotoxin AspF1 and siderophores was synthesized and Activated by HapX factor
Regulation
Regulates iron activating SreA function by post translationally.
Regulates iron blocking HapX function by post translationally.
inactivation of either genes leads to
During iron sufficiency and/or starvation, SreA are confined to growth with their mode of action and expression pattern.
Represses excessive uptake of iron through the down regulation of siderophore mediated and reductive iron assimilation, under iron replete conditions and retardation of growth this result in iron toxic accumulation.
During iron sufficiency and/or deficiency, HapX are confined to growth with their mode of action and expression pattern and reduced TAFC production.
6. Iron acquisition and storage in A. fumigatus:
For host iron sources, A. fumigatus lacks particular uptake systems such as heme, Transferring or ferritin. To mobilize extracellular iron, it excretes fusarinine C (FsC) and triacetylfusarinine C (TAFC) (Table 5). Following to iron chelation, the ferric forms (fe III) of Fs C and TAFC are taken up by Specific Iron Transporters (SIT) (Haas., et al., 2003) and for storage and distribution of iron consists of two different siderophores excretes intracellularly such as hyphal ferricrocin (FC) and conidial hydroxyferricrocin (HFC) shown in (Table 5) (Schrettl., et al., 2007 and Wallner., et al., 2009).
For release of iron, the siderophores are intracellularly hydrolyzed (Almeida., et al., 2009) and the iron is transferred or stored to the metabolic machinery. In fungi, four types of different mechanisms for iron acquisition have been identified, they are: (Haas., et al., 2008).
Uptake of specific iron chelators of ferric iron known as siderophores
low-affinity ferrous iron uptake
uptake of heme and degradation
Reductive Iron Assimilation (RIA)
SIT constitutes a subfamily of the major facilitator superfamily of protein acting most probably vitalized by the plasma membrane potential as proton symporters Philpott and Protchenko., 2008) [67]. In the fungal kingdom, SIT attributes uptake of iron visualizes to be protected universally [68, 69]. Iron supply is ensured by low affinity ferrous iron acquisition, reductive iron assimilation (RIA), two high affinity iron uptake systems, siderophore mediated iron uptake [70].
Additionally, A. fumigatus employs vacuolar iron storage and CccA expression indicated by the iron inducible system [59]. During infection, removal of iron from host sources such as transferring by iron acquisition plays major role in A. fumigatus siderophores [71,72].
The ferrous iron which is reduced is reoxidized and imported consisting of the ferroxidase FetC and the iron permease FtrA by a protein complex. RIA starts with ferriciron reduction to the high soluble ferrous iron by plasma membrane localized metallo reductases [73]. A. fumigatus encodes putative metalloreductases FreB has involved in Reductive Irion Assimilation indicating possible enzyme dismissal. Blocking of this RIA (Δ ftrA mutant) does not have any effect of A. fumigatus virulence.
In order to overcome the stages of iron toxicity and iron limiting conditions iron-limitation as well as to avoid iron toxicity (Table 3), iron should be stored in cells. To maintain as such there as two types of strategies involved:
Iron deposition in vacuoles and
siderophore-mediated iron storage [59] and [74]
For storage ofiron, A. fumigates engage in both vacuolar iron deposition and intracellular siderophores [75]
7. Iron transport mechanisms in A. fumigatus:
In general, iron transport channels for siderophore mediated have four different types of mechanisms and the fungal species involved in this transport are shown in (Table 7)[76]:
Shuttle mechanism
Taxicab mechanism
Hydrolytic mechanism
Reductive mechanism
Shuttle mechanism: From the ligand the ferrous oxide (Fe-III) is released and reduced by reductive enzymes and the siderophores which are free are then recycled, whereas siderophore complex- ferrous oxide is transported across the cell membrane [77].
Taxicab mechanism: Ferrous oxide (Fe-III) transfers cell membrane from the extracellular siderophores to intracellular ligands [78]
Hydrolytic mechanism: To release the ferrous oxide (Fe III) the complete Fe (III)–siderophore complex is transported into the cell which is related to degradative and several reductive process. Inside the cell, Ferrous oxide is reduced to Ferric oxide and then siderophore is again excreted [79].
Reductive mechanism: Siderophore complex of Fe (III) does not transport across the cell membrane. The reduced iron is taken up by the cell soon after the reduction of ferrous oxide to ferric oxide at the cell membranes [80].
Table 7: Iron transport mechanisms in fungal siderophores
S.no
Mechanisms
Fungal sps.
Transporting fungal siderophores
Refereneces
1
shuttle mechanism
Ustilago maydis
Ferrichrome
Ardon et al., (1998) [77]
2
taxicab mechanism
Rhodotorula species
Winkelmann and Huschka., (1987) [78]
3
hydrolytic mechanism
Mycelia sterilia
Adjimani and Emery., (1988) [79]
4
reductive mechanism
Ustilago
Sphaerogena
Ferrichrome
Ecker and Emery., (1983) [80]
(Ahmad and Holmostrom 2014)
8.PATENTS:
Patents granted on the detection of siderophores in Aspergillus fumigatus were listed below in table 8
Table 8: List Of Patents Granted on Detection of A. fumigatus
Patents list
Patent number
Date of patent published
Assignee
Inventors
Aspergillus fumigatus lectin and methods of using same
WO1997044061A1
1997-11-27
Toshiyuki Ishimaru,
Edward M. Bernard,
Donald Armstrong,
David A. Sheinberg,
Monica Bocchia,
Maurice E. Murphy
Nucleic acid probes for the detection and identification of fungi
Production of mutant strain of Aspergillus fumigatus, method of assay for inhibiting siderophore biosynthesis and diagnostic method for detecting likely Aspergillus fumigatus infection
The USA, represented by The Department of Health and Human Services
Production of mutant strain of Aspergillus fumigatus, method of assay for inhibiting siderophore biosynthesis and diagnostic method for detecting likely Aspergillus fumigatus infection
Council of Scientific and Industrial Research (CSIR)
Preetida Jagdish Bhetariya,
Taruna Madan Gupta,
Yogendra Singh,
Anupam Varma,
Puranam Usha Sarma.
Methods and compositions for the inhibition of growth of infectious Aspergillus fumigatus and other mycotic organisms in the gut of mammalian and avian species
A. fumigatus saprophytic fungi play an unpleasant role in human organs as a invasive aspergillosis. The both extracellular and intracellular siderophores togetherly plays as an important function of A. fumigatus virulence in a invasive pulmonary aspergillosis of murine model, but blocking of metalloreductase FreB mutant involves in RIA for the great affinity manner to acquire iron.
Srb A was found to be active siderophore, as this deficiency leads to decrease in ergosterol content. Furthermore, this leads to low level of resistance in against azole drugs. Therefore, ergosterol biosynthetic pathway should be highly improved for the best diagnosis of fungal infections hostile to humans and through this mortality rate can also be low.
In general, fungal siderophores transports Iron uptake in different mechanisms for the absorption and regulation of ferrous form of ions (feII), that may be predictable in A. fumigatus in future perspectives.
Funding: No funding sources
Conflict of interest: None declared
Ethical approval: The study was approved by the Institutional Ethics Committee of Biological Sciences, Allahabad
References
Kwon-Chung, Kyung J., and Janyce A. Sugui. "Aspergillus fumigatus—what makes the species a ubiquitous human fungal pathogen?." PLoS pathogens 9.12 (2013): e1003743.
Hirsch, T., et al. "House‐dust‐mite allergen concentrations (Der f 1) and mold spores in apartment bedrooms before and after installation of insulated windows and central heating systems." Allergy 55.1 (2000): 79-83.
Beffa, T., et al. "Mycological control and surveillance of biological waste and compost." Medical Mycology 36.1 (1998): 137-145.
Ryckeboer, Jaak, et al. "Microbiological aspects of biowaste during composting in a monitored compost bin." Journal of Applied microbiology 94.1 (2003): 127-137.
Dixon, D. D., and T. J. Walsh. "Aspergillus, biology and industrial application." Journal of Human pathogenesis (1992): 249-267.
Kwon-Chung, K. J., & Bennett, J. E. (1992). Medical mycology. Lea & Febiger, Philadelphia, Pa.
Pennington J. E. (1988). Respiratory infections: diagnosis and management. Raven Press, New York.
Zureik, Mahmoud, et al. "Sensitisation to airborne moulds and severity of asthma: cross sectional study from European Community respiratory health survey." Bmj 325.7361 (2002): 411.
Grigoriv, D., Bubmule, J., & Delacretaz, J. (1979). Aspergillus sinusitis. Postgraduate Medical Journal. 55, 619–21.
Rudwan, M. A., and H. A. Sheikh. "Aspergilloma of paranasal sinuses—a common cause of unilateral proptosis in Sudan." Clinical Radiology 27.4 (1976): 497-502.
Rothfeld, I., CF BEGG, and G. ELIOT. "Aspergilloma of sinus." New York State Journal of Medicine 72.4 (1972): 493-+.
Rosenberg, Michael, et al. "Clinical and immunologic criteria for the diagnosis of allergic bronchopulmonary aspergillosis." Annals of Internal Medicine 86.4 (1977): 405-414.
Agarwal, R., et al. "Allergic bronchopulmonary aspergillosis: review of literature and proposal of new diagnostic and classification criteria." Clinical & Experimental Allergy 43.8 (2013): 850-873.
Wald, Anna, et al. "Epidemiology of Aspergillus infections in a large cohort of patients undergoing bone marrow transplantation." The Journal of infectious diseases 175.6 (1997): 1459-1466.
Holland, Steven M. "Chronic granulomatous disease." Clinical reviews in allergy & immunology 38 (2010): 3-10.
De Bock, R. "Epidemiology of invasive fungal infections in bone marrow transplantation. EORTC Invasive Fungal Infections Cooperative Group." Bone Marrow Transplantation 14 (1994): S1-2.
Marr, Kieren A., Thomas Patterson, and David Denning. "Aspergillosis: pathogenesis, clinical manifestations, and therapy." Infectious Disease Clinics 16.4 (2002): 875-894.
Gifford, Anna HT, Jodine R. Klippenstein, and Margo M. Moore. "Serum stimulates growth of and proteinase secretion by Aspergillus fumigatus." Infection and immunity 70.1 (2002): 19-26.
d'Enfert, Christophe. "Fungal Spore Germination: Insights from the Molecular Genetics ofAspergillus nidulansandNeurospora crassa." Fungal genetics and Biology 21.2 (1997): 163-172.
Fontaine, Thierry, et al. "Molecular organization of the alkali-insoluble fraction ofAspergillus fumigatus cell wall." Journal of Biological Chemistry 275.36 (2000): 27594-27607.
Gardiner, Donald M., Renée S. Jarvis, and Barbara J. Howlett. "The ABC transporter gene in the sirodesmin biosynthetic gene cluster of Leptosphaeria maculans is not essential for sirodesmin production but facilitates self-protection." Fungal genetics and biology 42.3 (2005): 257-263.
Dufresne, Simon F., et al. "Detection of urinary excreted fungal galactomannan-like antigens for diagnosis of invasive aspergillosis." (2012): e42736.
Stephens-Romero, Shane D., Aron J. Mednick, and Marta Feldmesser. "The pathogenesis of fatal outcome in murine pulmonary aspergillosis depends on the neutrophil depletion strategy." Infection and immunity 73.1 (2005): 114-125.
Lehmann, P. F., and E. Reiss. "Invasive aspergillosis: antiserum for circulating antigen produced after immunization with serum from infected rabbits." Infection and immunity 20.2 (1978): 570-572.
Tekaia, Fredj, and Jean-Paul Latgé. "Aspergillus fumigatus: saprophyte or pathogen?." Current opinion in microbiology 8.4 (2005): 385-392.
Nierman, William C., et al. "Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus fumigatus." Nature 438.7071 (2005): 1151-1156.
Schrettl, Markus, et al. "Siderophore biosynthesis but not reductive iron assimilation is essential for Aspergillus fumigatus virulence." The Journal of experimental medicine 200.9 (2004): 1213-1219.
Kaplan, Craig D., and Jerry Kaplan. "Iron acquisition and transcriptional regulation." Chemical reviews 109.10 (2009): 4536-4552.
Gamit, D. A., and S. K. Tank. "Effect of siderophore producing microorganism on plant growth of Cajanus cajan (Pigeon pea)." Int J Res Pure Appl Microbiol 4.1 (2014): 20.
Bou-Abdallah, Fadi. "The iron redox and hydrolysis chemistry of the ferritins." Biochimica et Biophysica Acta (BBA)-General Subjects 1800.8 (2010): 719-731.
Taylor, Kevin G., and Kurt O. Konhauser. "Iron in earth surface systems: a major player in chemical and biological processes." Elements 7.2 (2011): 83-88.
Messenger, Ann JM, and Raymond Barclay. "Bacteria, iron and pathogenicity." Biochemical Education 11.2 (1983): 54-63.
Fardeau, Sylvain, et al. "Bacterial iron uptake: a promising solution against multidrug resistant bacteria." Science against microbial pathogens: communicating current research and technological advances 2 (2011): 695-705.
Hubertus Haas. (2012). Iron–a key nexus in the virulence of Aspergillus fumigatus. Frontiers in microbiology. 3, 1-10.
Halliwell, Barry, and JMe Gutteridge. "Oxygen toxicity, oxygen radicals, transition metals and disease." Biochemical journal 219.1 (1984): 1.
Simoes, Lúcia Chaves, Manuel Simoes, and Maria Joao Vieira. "Biofilm interactions between distinct bacterial genera isolated from drinking water." Applied and environmental microbiology 73.19 (2007): 6192-6200.
Blatzer, Michael, et al. "SidL, an Aspergillus fumigatus transacetylase involved in biosynthesis of the siderophores ferricrocin and hydroxyferricrocin." Applied and environmental microbiology 77.14 (2011): 4959-4966.
Schrettl, Markus, et al. "HapX-mediated adaption to iron starvation is crucial for virulence of Aspergillus fumigatus." PLoS pathogens 6.9 (2010): e1001124.
Schwyn, Bernhard, and J. B. Neilands. "Universal chemical assay for the detection and determination of siderophores." Analytical biochemistry 160.1 (1987): 47-56.
Nagoba, Basavraj, and Deepak V. VEDPATHAK. "Medical applications of siderophores–a review." European Journal of General Medicine 8.3 (2011): 230-233.
Oberegger, Harald, et al. "4′-Phosphopantetheinyl transferase-encoding npgA is essential for siderophore biosynthesis in Aspergillus nidulans." Current genetics 44 (2003): 211-215.
Allen, Gethin, et al. "Functional analysis of a mitochondrial phosphopantetheinyl transferase (PPTase) gene pptB in Aspergillus fumigatus." Fungal genetics and biology 48.4 (2011): 456-464.
Hossain, M. BILAYET, M. A. F. Jalal, and D. Van der Helm. "6-L-alanineferrirubin, a ferrichrome-type siderophore from the fungus Aspergillus ochraceous." Acta Crystallographica Section C: Crystal Structure Communications 53.6 (1997): 716-718.
Koulman, Albert, et al. "Identification of extracellular siderophores and a related peptide from the endophytic fungus Epichloë festucae in culture and endophyte-infected Lolium perenne." Phytochemistry 75 (2012): 128-139.
Yin, Wen-Bing, et al. "A nonribosomal peptide synthetase-derived iron (III) complex from the pathogenic fungus Aspergillus fumigatus." Journal of the American Chemical Society 135.6 (2013): 2064-2067.
Schrettl, Markus, et al. "HapX-mediated adaption to iron starvation is crucial for virulence of Aspergillus fumigatus." PLoS pathogens 6.9 (2010): e1001124.
Kragl, Claudia, et al. "EstB-mediated hydrolysis of the siderophore triacetylfusarinine C optimizes iron uptake of Aspergillus fumigatus." Eukaryotic cell 6.8 (2007): 1278-1285.
Blatzer, Michael, et al. "SidL, an Aspergillus fumigatus transacetylase involved in biosynthesis of the siderophores ferricrocin and hydroxyferricrocin." Applied and environmental microbiology 77.14 (2011): 4959-4966.
Winterberg, Britta, et al. "Elucidation of the complete ferrichrome A biosynthetic pathway in Ustilago maydis." Molecular microbiology 75.5 (2010): 1260-1271.
Yasmin, Sabiha, et al. "Mevalonate governs interdependency of ergosterol and siderophore biosyntheses in the fungal pathogen Aspergillus fumigatus." Proceedings of the National Academy of Sciences 109.8 (2012): E497-E504.
Schrettl, Markus, et al. "Siderophore biosynthesis but not reductive iron assimilation is essential for Aspergillus fumigatus virulence." The Journal of experimental medicine 200.9 (2004): 1213-1219.
Hissen, A. H., & Moore, M. M. (2005). Site specific rate constants for iron acquisition from transferring by the Aspergillu sfumigatus siderophores N_,N__,N__ _-triacetyl fusarinine C and ferricrocin. Journal of Biolpgical Inorganic Chemistry. 10, 211–220.
Haas, Hubertus. "Fungal siderophore metabolism with a focus on Aspergillus fumigatus." Natural product reports 31.10 (2014): 1266-1276.
Schrettl, Markus, et al. "Distinct roles for intra-and extracellular siderophores during Aspergillus fumigatus infection." PLoS pathogens 3.9 (2007): e128.
Kragl, Claudia, et al. "EstB-mediated hydrolysis of the siderophore triacetylfusarinine C optimizes iron uptake of Aspergillus fumigatus." Eukaryotic cell 6.8 (2007): 1278-1285.
Eisendle, Martin, et al. "The intracellular siderophore ferricrocin is involved in iron storage, oxidative-stress resistance, germination, and sexual development in Aspergillus nidulans." Eukaryotic Cell 5.10 (2006): 1596-1603.
Haas, Hubertus, et al. "The Aspergillus nidulans GATA factor SREA is involved in regulation of siderophore biosynthesis and control of iron uptake." Journal of Biological Chemistry 274.8 (1999): 4613-4619.
Hortschansky, Peter, et al. "Interaction of HapX with the CCAAT‐binding complex—a novel mechanism of gene regulation by iron." The EMBO journal 26.13 (2007): 3157-3168.
Schrettl, Markus, et al. "SreA‐mediated iron regulation in Aspergillus fumigatus." Molecular microbiology 70.1 (2008): 27-43.
Schrettl, Markus, et al. "HapX-mediated adaption to iron starvation is crucial for virulence of Aspergillus fumigatus." PLoS pathogens 6.9 (2010): e1001124.
Willger, Sven D., et al. "A sterol-regulatory element binding protein is required for cell polarity, hypoxia adaptation, azole drug resistance, and virulence in Aspergillus fumigatus." PLoS pathogens 4.11 (2008): e1000200.
Simoes, Lúcia Chaves, Manuel Simoes, and Maria Joao Vieira. "Biofilm interactions between distinct bacterial genera isolated from drinking water." Applied and environmental microbiology 73.19 (2007): 6192-6200.
Bien, Clara M., and Peter J. Espenshade. "Sterol regulatory element binding proteins in fungi: hypoxic transcription factors linked to pathogenesis." Eukaryotic cell 9.3 (2010): 352-359.
Blatzer, Michael, Ulrike Binder, and Hubertus Haas. "The metalloreductase FreB is involved in adaptation of Aspergillus fumigatus to iron starvation." Fungal genetics and biology 48.11 (2011): 1027-1033.
Oberegger, Harald, et al. "SREA is involved in regulation of siderophore biosynthesis, utilization and uptake in Aspergillus nidulans." Molecular microbiology 41.5 (2001): 1077-1089.
Jain, Radhika, et al. "The MAP kinase MpkA controls cell wall integrity, oxidative stress response, gliotoxin production and iron adaptation in Aspergillus fumigatus." Molecular microbiology 82.1 (2011): 39-53.
Philpott, Caroline C., and Olga Protchenko. "Response to iron deprivation in Saccharomyces cerevisiae." Eukaryotic cell 7.1 (2008): 20-27.
Schrettl, Markus, Günther Winkelmann, and Hubertus Haas. "Ferrichrome in Schizosaccharomyces pombe–an iron transport and iron storage compound." Biometals 17 (2004): 647-654.
Jung, Won Hee, and James W. Kronstad. "Iron and fungal pathogenesis: a case study with Cryptococcus neoformans." Cellular microbiology 10.2 (2008): 277-284.
Schrettl, Markus, Günther Winkelmann, and Hubertus Haas. "Ferrichrome in Schizosaccharomyces pombe–an iron transport and iron storage compound." Biometals 17 (2004): 647-654.
Hissen, A. H. T., et al. "Survival of Aspergillus fumigatus in serum involves removal of iron from transferrin: the role of siderophores." Infection and immunity 72.3 (2004): 1402-1408.
Hissen, A. H., & Moore, M. M. (2005). Site specific rate constants for iron acquisition from transferring by the Aspergillu sfumigatus siderophores N_,N__,N__ _-triacetyl fusarinine C and ferricrocin. Journal of Biolpgical Inorganic Chemistry. 10, 211–220.
Kosman, Daniel J. "Redox cycling in iron uptake, efflux, and trafficking." Journal of Biological Chemistry 285.35 (2010): 26729-26735.
Kaplan, Craig D., and Jerry Kaplan. "Iron acquisition and transcriptional regulation." Chemical reviews 109.10 (2009): 4536-4552.
Gsaller, Fabio, et al. "The interplay between vacuolar and siderophore-mediated iron storage in Aspergillus fumigatus." Metallomics 4.12 (2012): 1262-1270.
van der Helm, Dick, and Günther Winkelmann. "Hydroxamates and polycarboxylates as iron transport agents (siderophores) in fungi." Metal ions in fungi. CRC Press, 2020. 39-98.
Ardon, Orly, et al. "Iron uptake in Ustilago maydis: tracking the iron path." Journal of bacteriology 180.8 (1998): 2021-2026.
Winkelmann, Günther, and Hans-Georg Huschka. "Molecular recognition and transport of siderophores in fungi." Iron transport in microbes, plants and animals/edited by Gunther Winkelmann, Dick van der Helm, Joe B. Neilands (1987).
Adjimani, JONATHAN P., and T. H. O. M. A. S. Emery. "Stereochemical aspects of iron transport in Mycelia sterilia EP-76." Journal of bacteriology 170.3 (1988): 1377-1379.
Ecker, DAVID J., and T. H. O. M. A. S. Emery. "Iron uptake from ferrichrome A and iron citrate in Ustilago sphaerogena." Journal of bacteriology 155.2 (1983): 616-622.
Advertisement
Recommended Articles
Research Article
Study of Marination of Broiler Chicken with Garlic on Water Holding City, Cooking Loss, Tenderness and Number of Bacteria
Lilis Suryaningsih,
...
Dicki Tri Utama
Published: 20/06/2025
Download PDF
Cite
x
APA
Suryaningsih, L., Gumilar, J., Setyadi Putranto, W., Wulandari, E., None, P. & Tri Utama, D. (2025). Study of Marination of Broiler Chicken with Garlic on Water Holding City, Cooking Loss, Tenderness and Number of Bacteria. Himalayan Journal of Agriculture, 6(1), 1-4.
MLA
Suryaningsih, Lilis, et al. "Study of Marination of Broiler Chicken with Garlic on Water Holding City, Cooking Loss, Tenderness and Number of Bacteria." Himalayan Journal of Agriculture 6.1 (2025): 1-4.
Chicago
Suryaningsih, Lilis, Jajang Gumilar, Wendry Setyadi Putranto, Eka Wulandari, Pratama and Dicki Tri Utama. "Study of Marination of Broiler Chicken with Garlic on Water Holding City, Cooking Loss, Tenderness and Number of Bacteria." Himalayan Journal of Agriculture 6, no. 1 (2025): 1-4.
Harvard
Suryaningsih, L., Gumilar, J., Setyadi Putranto, W., Wulandari, E., None, P. and Tri Utama, D. (2025) 'Study of Marination of Broiler Chicken with Garlic on Water Holding City, Cooking Loss, Tenderness and Number of Bacteria' Himalayan Journal of Agriculture 6(1), pp. 1-4.
Vancouver
Suryaningsih L, Gumilar J, Setyadi Putranto W, Wulandari E, Pratama P, Tri Utama D. Study of Marination of Broiler Chicken with Garlic on Water Holding City, Cooking Loss, Tenderness and Number of Bacteria. Himalayan Journal of Agriculture. 2025 Jan;6(1):1-4.
Download PDF
Research Article
Mating behaviour of Cavariella aegopodii
Nidhi Dhatwalia
Published: 19/07/2024
Download PDF
Cite
x
APA
Dhatwalia, N. (2024). Mating behaviour of Cavariella aegopodii. Himalayan Journal of Agriculture, 5(1), 1-7.
MLA
Dhatwalia, Nidhi. "Mating behaviour of Cavariella aegopodii." Himalayan Journal of Agriculture 5.1 (2024): 1-7.
Chicago
Dhatwalia, Nidhi. "Mating behaviour of Cavariella aegopodii." Himalayan Journal of Agriculture 5, no. 1 (2024): 1-7.
Harvard
Dhatwalia, N. (2024) 'Mating behaviour of Cavariella aegopodii' Himalayan Journal of Agriculture 5(1), pp. 1-7.
Vancouver
Dhatwalia N. Mating behaviour of Cavariella aegopodii. Himalayan Journal of Agriculture. 2024 Jan;5(1):1-7.
Download PDF
Research Article
The Effect of Marination Duration using Tamarind Paste (Tamarindus indica L.) on pH, Tenderness, Water-holding Capacity and Cooking Loss of Spent Layer Meat
Dina Ayu Wulandari,
...
Dicky Tri Utama
Published: 30/06/2024
Download PDF
Cite
x
APA
Wulandari, D. A., Suryaningsih, L. & Utama, D. T. (2024). The Effect of Marination Duration using Tamarind Paste (Tamarindus indica L.) on pH, Tenderness, Water-holding Capacity and Cooking Loss of Spent Layer Meat. Himalayan Journal of Agriculture, 5(1), 1-12.
MLA
Wulandari, Dina A., Lilis Suryaningsih and Dicky T. Utama. "The Effect of Marination Duration using Tamarind Paste (Tamarindus indica L.) on pH, Tenderness, Water-holding Capacity and Cooking Loss of Spent Layer Meat." Himalayan Journal of Agriculture 5.1 (2024): 1-12.
Chicago
Wulandari, Dina A., Lilis Suryaningsih and Dicky T. Utama. "The Effect of Marination Duration using Tamarind Paste (Tamarindus indica L.) on pH, Tenderness, Water-holding Capacity and Cooking Loss of Spent Layer Meat." Himalayan Journal of Agriculture 5, no. 1 (2024): 1-12.
Harvard
Wulandari, D. A., Suryaningsih, L. and Utama, D. T. (2024) 'The Effect of Marination Duration using Tamarind Paste (Tamarindus indica L.) on pH, Tenderness, Water-holding Capacity and Cooking Loss of Spent Layer Meat' Himalayan Journal of Agriculture 5(1), pp. 1-12.
Vancouver
Wulandari DA, Suryaningsih L, Utama DT. The Effect of Marination Duration using Tamarind Paste (Tamarindus indica L.) on pH, Tenderness, Water-holding Capacity and Cooking Loss of Spent Layer Meat. Himalayan Journal of Agriculture. 2024 Jan;5(1):1-12.
Download PDF
Research Article
The Role of Indigenous Microbes and Earthworm in the Bioconversion of Dairy Wastewater Solids into Organic Fertilizer
Marlina ,
...
N. Meynadhea
Published: 17/01/2024
Download PDF
Cite
x
APA
None, M., Rahayu, N. A., Badruzzaman, D. Z., Hidayati, Y., Harlia, E., Kurniawan, E. & Meynadhea, N. (2024). The Role of Indigenous Microbes and Earthworm in the Bioconversion of Dairy Wastewater Solids into Organic Fertilizer. Himalayan Journal of Agriculture, 5(1), 1-7.
MLA
None, Marlina, et al. "The Role of Indigenous Microbes and Earthworm in the Bioconversion of Dairy Wastewater Solids into Organic Fertilizer." Himalayan Journal of Agriculture 5.1 (2024): 1-7.
Chicago
None, Marlina, N. A. Rahayu, D. Z. Badruzzaman, Y.A. Hidayati, E. Harlia, E. Kurniawan and N. Meynadhea. "The Role of Indigenous Microbes and Earthworm in the Bioconversion of Dairy Wastewater Solids into Organic Fertilizer." Himalayan Journal of Agriculture 5, no. 1 (2024): 1-7.
Harvard
None, M., Rahayu, N. A., Badruzzaman, D. Z., Hidayati, Y., Harlia, E., Kurniawan, E. and Meynadhea, N. (2024) 'The Role of Indigenous Microbes and Earthworm in the Bioconversion of Dairy Wastewater Solids into Organic Fertilizer' Himalayan Journal of Agriculture 5(1), pp. 1-7.
Vancouver
Marlina M, Rahayu NA, Badruzzaman DZ, Hidayati Y, Harlia E, Kurniawan E, Meynadhea N. The Role of Indigenous Microbes and Earthworm in the Bioconversion of Dairy Wastewater Solids into Organic Fertilizer. Himalayan Journal of Agriculture. 2024 Jan;5(1):1-7.
Kumeera, B., John, S. A., Matikhaye, S. & Ramteke, P. W. (2021). Siderophores in Aspergillus Fumigatus: Biosynthesis and Iron Transport. Himalayan Journal of Agriculture, 2(1), 1-15.
MLA
Kumeera, Bethala, et al. "Siderophores in Aspergillus Fumigatus: Biosynthesis and Iron Transport." Himalayan Journal of Agriculture 2.1 (2021): 1-15.
Chicago
Kumeera, Bethala, Suchit A. John, Swapnil Matikhaye and Pramod W. Ramteke. "Siderophores in Aspergillus Fumigatus: Biosynthesis and Iron Transport." Himalayan Journal of Agriculture 2, no. 1 (2021): 1-15.
Harvard
Kumeera, B., John, S. A., Matikhaye, S. and Ramteke, P. W. (2021) 'Siderophores in Aspergillus Fumigatus: Biosynthesis and Iron Transport' Himalayan Journal of Agriculture 2(1), pp. 1-15.
Vancouver
Kumeera B, John SA, Matikhaye S, Ramteke PW. Siderophores in Aspergillus Fumigatus: Biosynthesis and Iron Transport. Himalayan Journal of Agriculture. 2021 Jan;2(1):1-15.