The major cause of dental caries is attributed to Streptococcus mutans. Dental plaque which also known as oral biofilm that forms when sugar is present, S. mutans is stimulated to secrete insoluble glucans which leads to the initiation of caries lesion. Compared with synthetic antibiofilm agents, natural antibiofilm agents such as herbal plants have few adverse effects and great economic value. The purpose of this paper is to study the antimicrobial effectiveness of cinnamaldehyde, a natural plant compound, against biofilms and virulence factors of S.mutans. We have demonstrated the effects of the antimicrobial of Cinnamaldehyde on S. mutans was evaluated by measuring the MIC and MBC, growth curves, acid production, and acid tolerance. Biofilm formation by S.mutans in the presence of sub-MIC concentrations of cinnamaldehyde was measured using a crystal violet assay, and virulence-related gene expression (gtfB, gtfC, gtfD, gbpB, and LuxS) was quantified using real-time PCR. The result have shown that S. mutans was inhibited by cinnamaldehyde with a MIC of 800 µg/mL and an MBC of 1600 µg/mL, the sub-MIC concentrations of cinnamaldehyde inhibited acid production and acid tolerance. In the presence of cinnamaldehyde, genes related to biofilms and acid tolerance were also downregulated. Therefore, we hypothesis that cinnamaldehyde possesses antimicrobial properties that effectively inhibit the growth of S. mutans by targeting various key aspects of its pathogenicity, including “biofilm formation, acid production, acid tolerance, and virulence gene expression.” This study will provide the optimization of preventing and therapeutic strategies for dental caries.
Keywords
Streptococcus mutans
Acid tolerance
Biofilm
Cinnamaldehyde.
INTRODUCTION
Dental caries is a widespread disease affecting a large portion of the global population, it is primarily caused by the presence of biofilm. Extensive research indicates that more than 90% of individuals worldwide are affected by dental caries [1, 2, 5, 6]. Biofilm refers to a three-dimensional structure consisting of various microorganisms residing in the oral cavity. If left untreated or not intervened with for an extended period the biofilm can mature and lead to the development of dental caries. Microorganisms within a biofilm particularly those contained an exopolysaccharide matrix bearing greater harm due to their increased resistance to the host immune system and different antimicrobial treatments [2, 3].
One of the major contributors to this critical condition is Streptococcus Mutans, which plays a significant role in forming complex and multidimensional structures on the oral mucosa and tooth enamel [1, 2, 4]. It acquires cariogenic properties, including the ability to adhere to solid surfaces, colonize the oral cavity and withstand the acidic environment within it [3]. Additionally, S. Mutans utilizes carbohydrates to produce acidic metabolites that lead to the degradation and demineralization of tooth enamel resulting in the reduction of mineral materials and the occurrence of dental caries [4].
Among the virulence factors of S. Mutans that contribute to the formation of oral biofilms are the Glucosyltransferases enzymes encoded by the gtfB, gtfC andgtfC genes [5]. These enzymes play a crucial role in producing [1-3] or [1-6] linked glucan polymers which are responsible for the formation of dental plaque [6, 7]. The maturation of dental plaque which contains polymicrobial communities requires cell-to-cell communication both within and between species. “S. Mutans also possesses a LuxS gene-mediated signaling pathway that influences biofilm formation, maturation and bacteriocin production [7, 8]”. This LuxS gene catalyzes the production of “autoinducer 2 (AI-2)”, a universally recognized signal that facilitates interspecies interactions in multispecies biofilms like dental plaque [9]. The S. Mutans LuxS gene is play an important role in various cellular activities such as biofilm formation and acid tolerance [7].
Controlling microbial accumulation (biofilm/plaque) on hard and soft surfaces of the mouth with natural cleaning (mastication), mechanical cleaning (brushing and flossing) and chemical plaque control (fissure sealant, fluoride supplement, sugar substitution and antimicrobial supplements) is one of the most effective oral health promotion strategies [10,11]. However, several antimicrobial drugs have been documented to have side effects with prolonged use [12]. These include tooth discoloration, poor taste and the induction of antimicrobial resistance strain. Culinary herbs, particularly those with medicinal properties (anti-inflammatory, antimicrobial) have gained popularity despite the fact that their safet, bioavailability and pharmacokinetic interactions have not been thoroughly studied [13].
Essential oil of cinnamon contains cinnamaldehyde, an α, β -unsaturated aromatic aldehyde [14]. It's a common additive to beverages, foods and fragrances [15]. Biofilms formed by Gram-positive and Gram-negative bacteria such as Pseudomonas aeruginosa and Staphylococcus aureus [14], have been shown to be susceptible to cinnamonaldehyde's antimicrobial effects. Therefore, we concentrated in this study on the antimicrobial activity of cinnamaldehyde at sub-minimum inhibitory concentration (sub-MIC) levels against S. Mutans biofilm. This research holds the potential to contribute towards the development of a natural product that could serve as a novel antivirulence agent targeting S. Mutans.
MATERIALS AND METHODS
S. Mutans
It was previously isolated from dental plaque in the microbiology lab of the university of wasit's college of medicine [16]. S. Mutans was cultured and maintained under optimal growth conditions in Brain Heart Infusion (BHI) broth.
Spectrophotometer
it is an instrument (UV1601, Shimadzu, Japan) that measure the number of photons absorbed after it passes through sample solution
Minimum Inhibitory Concentration and Minimum Bactericidal Concentration (MIC and MBC)
The protocol of the Clinical and Laboratory Standards Institute [17] broth method was used to determine the MIC and MBC of cinnamaldehyde against planktonic S. Mutans. Cinnamaldehyde (Sigma-Aldrich) was diluted in 0.5% Dimethyl Sulfoxide (DMSO) and tested at concentrations ranging from 200 to 2600 µg/mL. 0.5 McFarland of bacterial suspensions were added, followed by a 24-h incubation at 37°C. To ensure the validity of the methodology utilized in this study two control groups (a blank control consisting of sterile culture medium without cinnamaldehyde and microorganism suspensions and a vehicle control comprising sterile culture medium containing DMSO) were used as controls. These control groups were employed to account for any potential effects caused solely by the culture medium or the vehicle used in the experimental setup. The MIC of the cinnamaldehyde was the lowest concentration that prevented observable bacterial growth while the MBC was determined to be the lowest concentration that did not result in the growth of any colony when it was subcultured on agar plates. All experiments were performed in Triplicate and the results correspond to three independent experiments.
Growth Curve Assay
S. Mutans was grown anaerobically at 37 C° for 24 h in BHI media with varying doses of cinnamaldehyde (0, 50, 100, 200, 400 and 800 µg/mL). As a positive control, we employed chlorhexidine (0.2% concentration). Every 3 hours during incubation, the OD600nm was measured using a “spectrophotometer (UV1601, Shimadzu, Japan. Triplicate samples were used for each experiment and the results correspond to three independent experiments.
MTP-based Quantitative Biofilm Formation Test.
S. Mutans cultures at a concentration equivalent to 0.5 McFarland were diluted 100-fold in brain heart infusion broth containing 2% sucrose. Each diluted culture of the test isolates was dispensed into the wells of a “96-flat-bottom microtiter plate”, with 200 μl per well (3 wells per isolate). The microtiter plates were then subjected to anaerobic incubation at 37°C for 48 hours. Next the incubation period the plates underwent the following processing steps: The microtiter plates were inverted to drain the bacterial culture and then washed three to four times with sterile saline solution. After the washes, 200 µl of methanol was added to each well and the plates were left at room temperature for 20 minutes. The fixative methanol was then decanted and the plates were allowed to dry.
Next, the plates were stained by adding 200 μl of a 0.1% crystal violet solution to each well and incubating for 15 minutes. Afterward, the plates were washed three times with distilled water and allowed to dry by inverting them at room temperature. To solubilize the adhering cells and biofilms 200 μl of 33% glacial acetic acid was added to each well.
Using a microplate reader, the Optical Density (OD) of the samples was measured at 600 nm. Each clinical isolate was examined in triplicate the data were averaged and the standard deviation was computed. To account for background absorbance, the mean OD value of the control wells (which contained uninoculated broth) was subtracted from the average OD values of each test isolate [18]. The resulting values were then used to classify the test isolates based on their biofilm formation capabilities as presented in Table (1).
Table 1: Biofilm formation scales used to categorize S. Mutans isolates
OD value
Biofilm formation
OD≤2* ODc a
Weak biofilm producer
2*ODc≤OD≤4*ODc
Moderate biofilm producer
4*ODc≤OD
Strong biofilm producer
ODc is OD of the control wells
Anti-Biofilm Activity of Cinnamaldehyde
The experimental procedure followed the aforementioned steps, with the addition of the following details. Aliquots of 100 µl of cinnamaldehyde at a concentration of (0.5×MIC) were transferred to the first well of each row. A two-fold serial dilution was then performed, progressing up to the tenth column, after which 100 µl aliquots were discarded. The 11th and 12th columns served as positive controls, with the 11th column representing untreated isolates and the 12th column representing isolates treated with 0.2% chlorhexidine.
To assess the percent inhibition of biofilm formation caused by cinnamaldehyde for each test isolate, the following formula was used: [1– (average OD600 of treated isolate/average OD600 of untreated isolate)] × 100. “The Minimum Biofilm Inhibitory Concentration (MBIC) was determined by identifying the lowest concentration of the agent used that resulted in an inhibition of biofilm formation of 80% or more” [18].
Acidogenesis of S. Mutans.
The impact of cinnamaldehyde on S. Mutans glycolysis was determined according to [19], with minor modifications. Briefly, mid-logarithmic phase S. Mutans was collected, inoculated into 10 ml of tryptic soy broth (TSB, pH 7.5) and then resuspended in the same broth containing 50, 100, 200 and 400 g/mL of cinnamaldehyde. Over a period of 120 minutes the decrease in pH caused by the glycolytic activity of S. Mutans was monitored at 10-minute intervals and the experiment was repeated three times independently.
Acid tolerance assay of S. Mutans.
To assess acid tolerance, a broth dilution technique was employed. A series of Tryptic Soy Broth (TSB) with pH values ranging from 4.0 to 7.0, with intervals of 0.5, was prepared using the two-fold serial dilution technique. Cinnamaldehyde, on the other hand, was diluted in TPY liquid medium. For each sample, three replicates of each Cinnamaldehyde concentration were used, while three replicates without Cinnamaldehyde were employed as control groups. Subsequently, each tube was inoculated with a 1:10 (v/v) bacterial suspension under anaerobic conditions and “incubated at 37°C for 48 h”. After centrifuging the cultures for 15 minutes at 3000 r/min, the resulting bacterial cells were collected and washed three times with sterile deionized water. The optical density of the cell suspensions was measured at 600 nm using a spectrophotometer [20].
Effect of Cinnamaldehyde on Virulence Genes of S. Mutans
The impact of cinnamaldehyde on the expression of biofilm-related genes (luxS, gtfB, gtfC and gtfD) in S. Mutans was investigated using quantitative reverse transcription real-time PCR. The method described in [22] was followed, with the treated and control groups as outlined below:
Treatment S. Mutans isolates groups: S. Mutans was incubated in Tryptic Soy Broth (TSB) supplemented with 1% glucose and cinnamaldehyde at a concentration equal 100, 200 and 400 µg/ML. The incubation was conducted under anaerobic conditions at 37°C for 24 h
Control S. Mutans isolates groups: S. Mutans was incubated in TSB with 1% glucose alone, anaerobically at 37°C for 24 h
“Total RNA was extracted from the S. Mutans experimental isolates of the treatment and control groups using the TRIzol®” reagent kit, following the instructions provided by the manufacturer. The extracted total RNA was quantified and its purity was assessed using a “Nanodrop spectrophotometer (THERMO, USA)”.
To eliminate any residual genomic DNA, the extracted RNA samples were treated with DNase enzyme using a DNase I enzyme kit, following the protocol provided by the Promega company, USA. The inactivation of the DNase enzyme was carried out by adding EDTA to the mixture and incubating it at 37°C for 30 minutes.
The DNase-treated RNA samples were utilized for cDNA synthesis using the “AccuPower® RocktScript RT PreMix kit”.
The S. Mutans 16SrRNA gene was employed as an internal control for normalizing the expression of the target genes. Specific primers for luxS, gtfB, gtfC and gtfD genes of S. Mutans (as indicated in Table 2) were used. “Real-time PCR amplification was performed using a Real-time PCR thermocycler (Agilent, USA). Each reaction mixture of 25 μl contained 12.5 μl of 2x SYBR Green PCR Mix (TaKaRa, Japan), 1 μl of each primer (20 μM), 0.4 μl of ROX reference dye, 1 μl of sample cDNA and 9.1 μl of sterile deionized water. The amplification protocol included an initial denaturation step at 95°C for 2 minutes, followed by 40 cycles of denaturation at 95°C for 30 seconds, annealing at 55°C for 1 minute” and extension at 72°C for 1 minute. “All samples were run in triplicate and the relative quantification was performed using the 2-∆∆Ct method”. Each experiment included triplicate samples and the results represent three independent experiments.
RESULTS
Cinnamaldehyde's Antibacterial Effect on Planktonic S. Mutans
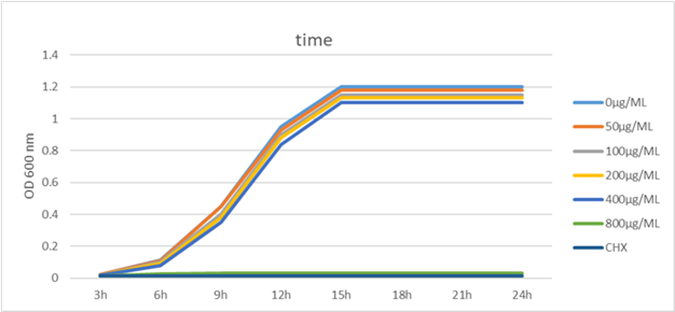
Cinnamaldehyde demonstrated a Minimum Inhibitory Concentration (MIC) of 800 µg/mL and a minimum bactericidal concentration (MBC) of 1600 µg/mL against planktonic S. Mutans. The growth curve assay results provided evidence that cinnamaldehyde effectively suppressed the growth of S. Mutans at a concentration of 800 µg/mL. In comparison to the control group, concentrations 400 µg/mL of cinnamaldehyde did not significantly alter the growth curve of S. Mutans, as depicted in Figure (1).
Table 2: Primers used in this study and their respective nucleotide sequences
Gene
Primer sequence (5’-3’)
Size (bp)
References
16S rRNA
“F: CCTACGGGAGGCAGCAG”
196
20
“R:ATTACCGCGGCTGCTGG”
luxS
“F: ACTTGCTTTGATGACTGTGGC”
115
20
“R:TCAGCGTATTGACGGGATG”
gtfB
“F: AGCAATGCAGCCAATCTACAAAT”
59
21
“R: ACGAACTTTGCCGTTATTGTCA”
gtfC
“F: GTGCGCTACACCAATGACAGAG”
107
21
“R: GCCTACTGGAACCCAAACACCTA”
gtfD
“F: TGGCACCGCAATATGTCTCTTC”
103
21
“R: CAATCCGCAATAACCTGAATACCG”
Figure 1: S. Mutans growth curve at various cinnamaldehyde concentration
Figure 2: Inhibition of S. Mutans acid production by cinnamaldehyde. “∗P<0.05, significantly different from the control group
Effect of Cinnamaldehyde on Acid Production S. Mutans
In order to assess the impact of “cinnamaldehyde on acid production by S. Mutans”, a glycolytic pH drop assay was conducted. S. Mutans cultures were exposed to different concentrations of cinnamaldehyde and the resulting pH changes were measured. The findings, as illustrated in Figure (2), indicate that in the control group, the pH decreased from 7.25±0.09 to 4.44±0.11 after 120 minutes of incubation. However, treatment with cinnamaldehyde at concentrations of 100, 200 and 400 µg/mL elevated the terminal pH from the acidic level (4.44±0.06) to 5.4±0.12, 5.62±0.09 and 6.1±0.13, respectively.
Furthermore, the control group exhibited the maximum initial pH drop within the first 20 minutes of incubation reducing from 7.25±0.09 to 4.91±0.16. In contrast, the concentration of 400 µg/mL cinnamaldehyde resulted in the minimum initial pH drop”, decreasing from 7.25±0.09 to 6.64±0.12. These observations suggest that cinnamaldehyde can mitigate the acid production by S. Mutans, as it reduced the extent of pH drop compared to the control group.
Effect of Cinnamaldehyde on Acidurity of S. Mutans.
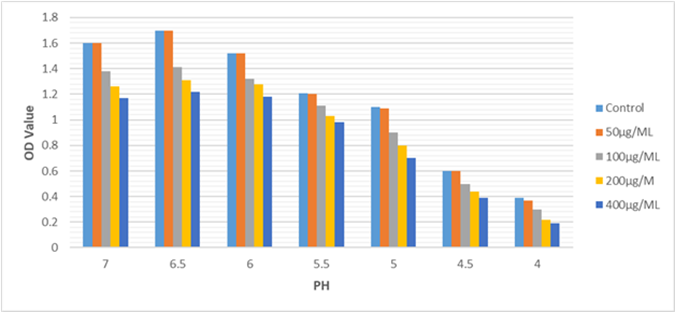
The impact of “Cinnamaldehyde on the S. Mutans acid tolerance was assessed and the results are presented in Figure (3).” Compared to the control groups, the 100 µg/mL, 200 µg/mL and 400 µg/mL -treated groups significantly inhibited the proliferation of S. Mutans at the same pH (P 0.05).
Figure 3: Effects of Cinnamaldehyde on acid tolerance of S. Mutans isolates at sub-MIC levels. The acid tolerance of S. Mutans was determined by measuring its optical density at 600 nm after 48 hours of treatment with Cinnamaldehyde. *statistically significant difference (P<0.05) in the groups that treated with 100µg/ML,200 µg/ML and 400 µg/ML compared with Control
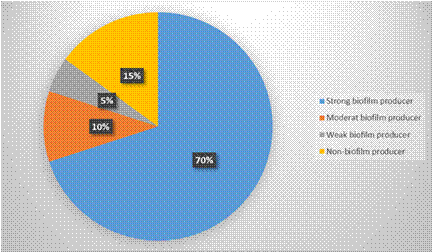
Figure 4: The proportion of S. Mutans isolates with “strong, moderate, and weak biofilm formation”
Moreover, “a dose-dependent inhibition” was observed across the pH range of 7.0 to 4. However, the concentration of 50 µg/mL did not demonstrate a significant effect on the growth of S. Mutans.
Detection of Biofilm Formation by Microtiter Plate (MTP) Assay
The assay results revealed that among the 20 S.Mutans isolates tested, 19 (95%) of them were capable of forming biofilm to varying degrees. Figure (4) illustrates that 14 (70%) isolates exhibited strong biofilm production, 2 (10%) isolates showed moderate biofilm production and 3 (15%) isolates displayed weak biofilm production. Only 1 (5%) isolate did not show any biofilm production.
Determination of Antibiofilm Activity of Cinnamaldehyde Against S. Mutans Isolates
Cinnamaldehyde was tested for antibiofilm activity using 19 S. Mutans isolates. As outlined in Table (3), the "Minimal Biofilm Inhibitory Concentration" (MBIC) of cinnamaldehyde against the test isolates ranged from 100 to 400 µg/ml.
Effects of Cinnamaldehyde on the expression of luxS, gtfB, gtfC and gtfD genes
Based on the results of the Microtiter Plate (MTP) assay, strong biofilms formation S. Mutans were selected for further analysis of biofilm-related gene expression using real-time PCR. The expression levels of luxS, gtfB, gtfC and gtfD genes in S. Mutans were examined and the findings are presented in Figure (5). Significant down-regulation of these genes was observed after treatment with 200 µg/mL and 400 µg/mL cinnamaldehyde compared to the control group. However, the concentration of 100 µg/mL cinnamaldehyde did not show any significant effects on gene expression.
Furthermore, with increasing cinnamaldehyde concentration, “relative fold changes of gene transcripts decreased”, as depicted in Figure 6. Following treatment with 200 µg/mL and 400 µg/mL cinnamaldehyde, the expression of luxS was notably reduced by approximately 7.8-fold and 11.53-fold, respectively.
Similarly, the expression levels of gtfB, gtfC and gtfD genes, which are associated with biofilm formation, were also decreased after cinnamaldehyde treatment.
Figure 6: Cinnamaldehyde's effect on the luxS, gtfB, gtfC, and gtfD gene expression. The results represent the average values and standard deviations obtained from three independent experiments conducted in triplicate. Statistical analysis revealed significant differences (“*P<0.05, **P<0.01”) between the experimental group and the control group
Table 3: Antibioflm activity of Cinnamaldehyde
Biofilm formation degree
Accession numbers
MBIC (µg /ml) of Cinnamaldehyde
Strong
OR016527
400
OR016434
400
OR016520
400
OR030436
200
OR016522
400
OR030432
400
OR016525
400
OR016526
400
OR016433
400
OR030427
200
OR030428
400
OR030429
400
OR030430
200
Moderate
OR030431
400
OR016521
200
Weak
OR030434
100
OR030435
200
OR016523
200
Minimum biofilm-inhibiting concentration (MBIC) was used to express the activity
At 200 µg/mL concentration, the fold changes were decreased in 2.72-fold (gtfB), 2.08-fold (gtfC) and 7.8-fold (gtfD). At a higher concentration of 400 µg/mL, the fold changes were decreased in 5.02-fold (gtfB), 4.28-fold (gtfC) and 16.66-fold (gtfD).
DISCUSSION
Dental caries is the most prevalent oral disease occurs due to a dysbiosis in tooth-associated to biofilms. Recent years have seen a growing interest in utilizing natural plant products to combat oral diseases leading to extensive research in this field [23]. In this particular study, we focused on cinnamaldehyde a natural product to investigate its antimicrobial properties against S. Mutans biofilm. Initially we determined the “MIC and MBC of cinnamaldehyde” against S. Mutans which were found to be 800 and 1600 µg/mL respectively. These values showed slight variations compared to previous studies. S. Mutans had a MIC of >500 M for cinnamaldehyde as determined by “resazurin staining with BHI broth at 37°C and 5% CO2 for 24 h” [24]. Additionally, the MBC value for S. Mutans was found to be 1728 µg/mL using TSB broth at 37°C for 24 hours [25]. The disparities in these results could be attributed to the use of different bacterial strains, culture media, conditions and methodologies. Especially, our findings indicated that concentrations of cinnamaldehyde below 400 µg/mL did not significantly impact the growth rate of the bacteria.
We study the influence of sub-MIC levels of cinnamaldehyde on the development of dental caries through the inhibition of S. Mutans biofilm formation. In the current study, CV assays demonstrated that increasing concentrations of cinnamaldehyde inhibited biofilm formation more effectively, this suggests that cinnamaldehyde has a dose-dependent effect on inhibiting the growth and development of S. Mutans biofilms. The increased effectiveness at higher concentrations indicates that cinnamaldehyde has a potent antimicrobial activity against S. Mutans, which is known to be a key contributor to the formation of dental caries.
The ability of S. Mutans to produce acid (acidogenicity) and withstand low pH levels (acidurance) are crucial physiological factors that contribute to the demineralization of tooth surfaces and the development of dental caries [5]. Using a glycolysis pH drop assay, we investigated the effect of sub-MIC concentrations of cinnamaldehyde on acid production in our study. Glycolysis is the primary acid production pathway, in which microorganisms metabolize diverse dietary carbohydrates to produce acid. The pH range of 5.0–5.5 is crucial because it determines the equilibrium between demineralization and remineralization of tooth enamel. When the ambient solution's pH falls below the critical pH due to acid accumulation, tooth demineralization and the onset of dental caries result [26].
Our findings demonstrated that as the concentration of cinnamaldehyde increased, the initial rate of pH decline decreased and the ultimate pH values exceeded the critical pH value. These findings suggest that cinnamaldehyde inhibits acidogenicity and prevents demineralization of teeth, possibly by inhibiting glycolytic enzymes responsible for acid production. The ability of microbes to tolerate acid is another significant factor associated with their cariogenic potential. Our research revealed that at pH 5.0, cinnamaldehyde reduces the survival rate of microorganisms. In addition, the ultimate pH values observed during the glycolytic pH drop assay are indicative of acid tolerance [27]. Thus, it is evident that cinnamaldehyde has a significant effect on “S. Mutans'acid production and acid tolerance”.
The effect of cinnamaldehyde on the expression of virulence factors in S. Mutans was subsequently evaluated using real-time PCR. Examining the variations in the expression of these genes revealed important information about their functions in biofilm formation and aided in the comprehension of the process. In the presence of cinnamaldehyde, the expression of all selection virulence genes was downregulated, according to our findings.
One of these genes is GTFase, it synthesizes glucans that serve as“binding sites for bacterial adhesion, biofilm formation and the development of dental caries. S. Mutans possesses at least three GTF enzymes (GTFB, GTFC and GTFD), each of which synthesizes a distinct form of glucan. GTFB, which is encoded by gtfB, generates water-insoluble polysaccharides containing 1,3-linked glucans and plays a crucial role in scaffolding the EPS matrix and promoting cell aggregation in stable biofilms. GTFC, which is encoded by gtfC, catalyzes the synthesis of a mixture of water-insoluble and alkali-soluble glucans from sucrose, incorporating both -1,3 and -6-linked glucans, which are required for plaque formation and structurally stable biofilms. GTFD, which is encoded by gtfD, synthesizes water-soluble glucans with -1,6-linked glucans, allowing interaction with salivary proteins in the pellicle” [28-30]. In vivo, mutant strains of S. Mutans with gtf gene mutations specifically gtfB and gtfC, have substantially reduced cariogenic potential compared to wild-type strains [31-33]. The downregulation of these gene expressions in our study suggests a reduction in the production of extracellular polysaccharides and oral bacterial aggregation thereby inhibiting biofilm formation.
Furthermore, cinnamaldehyde exhibited a reduction in the LuxS gene expression which encodes an enzyme responsible for catalyzing autoinducer 2(AI-2) production. AI-2 are crucial components of “inter- and intra-species microbial communication” in oral bacteria, including S. Mutans [7,9,34]. Therefore, the biofilm development of S. Mutans, which depends on interspecies communication was disrupted by downregulating the expression of luxS, which in turn prevented the synthesis of AI-2. Previous research has revealed that S. Mutans luxS mutants have a more granular appearance in biofilms and are unable to produce the AI-2 signal [7,8]. Moreover, it has been reported that luxS inactivation leads to a decrease in the” expression of several genes encoding membrane-associated proteins, which play roles in maintaining envelope integrity and acid tolerance” [35, 36]. Therefore, the downregulation of luxS gene expression observed in our study may have contributed to the decline in acid tolerance and biofilm formation.
Although this study provides evidence of the antimicrobial potential of cinnamaldehyde, there are still several mechanisms that require further investigation. Future research should focus on elucidating the effects of cinnamaldehyde on the structure and viability of monospecies and dual-species biofilms of the oral pathogens. Additionally, in vivo clinical trials and microbial analysis are necessary to establish the antimicrobial effects of cinnamaldehyde in a practical setting.
Statistical Analysis
The results are presented as the mean ± Standard Deviation (SD) (n = 3). Statistical processing of data was performed using Microsoft Excel 2010. All statistics were produced from at least three experiments using one-way ANOVA and the P value equal or less (P<0.05) was considered statistically significant difference.
CONCLUSION
Our study has demonstrated that cinnamaldehyde possesses antimicrobial properties that effectively inhibit the growth of S. Mutans by targeting various key aspects of its pathogenicity including biofilm formation, acid production, acid tolerance and virulence gene expression. These findings suggest that cinnamaldehyde has the potential to be used as a therapeutic agent against dental caries. However, further investigations are needed to fully understand the underlying mechanisms involved.
Future research should focus on exploring the specific effects of cinnamaldehyde on the structure and viability of both monospecies and dual-species biofilms. This will provide a more comprehensive understanding of its efficacy against complex microbial communities in the oral cavity. In addition, in vivo clinical trials and microbial analysis are necessary to evaluate the antimicrobial effects of cinnamaldehyde under realistic conditions and determine its potential for use in dental care.
Acknowledgment
We gratefully acknowledge the contribution of the Department of Microbiology, Wasit University College of Medicine, Iraq.
Conflict of Interest
There authors declare that they have no conflict of interest.
REFERENCES
Selwitz, R.H., et al. "Dental Caries." The Lancet, vol. 369, no. 9555, 2007, pp. 51–59.
Gurenlian, J.R. "American Dental Hygienists’ Association: The Role of Dental Plaque Biofilm in Oral Health." Journal of Dental Hygiene, vol. 81, no. 5, 2015, pp. 1–11.
Krzyściak, W., et al. The Virulence of Streptococcus Mutans and the Ability to Form Biofilms." European Journal of Clinical Microbiology & Infectious Diseases, vol. 33, no. 4, 2014, pp. 499–515. https://doi.org/10.1007/s10096-013-1993-7.
Ito, Y., et al. "Antimicrobial and Antibiofilm Effects of Abietic Acid on Cariogenic Streptococcus Mutans." Journal of Odontology, vol. 108, no. 1, 2020, pp. 57–65. https://doi.org/10.1007/s10266-019-00456-0.
Banas, J.A. "Virulence Properties of Streptococcus Mutans." Frontiers in Bioscience, vol. 9, 2004, pp. 1267–1277.
Burne, R.A. "Oral Streptococci: Products of Their Environment." Journal of Dental Research, vol. 77, no. 3, 1998, pp. 445–452.
Wen, Z.T. and R.A. Burne. "LuxS-Mediated Signaling in Streptococcus Mutans Is Involved in Regulation of Acid and Oxidative Stress Tolerance and Biofilm Formation." Journal of Bacteriology, vol. 186, no. 9, 2004, pp. 2682–2691.
Huang, Z., et al. "Lux S-Based Quorum Sensing Signaling Affects Biofilm Formation in Streptococcus Mutans." Journal of Molecular Microbiology and Biotechnology, vol. 17, no. 1, 2009, pp. 12–19.
Kolenbrander, P.E., et al. "Communication Among Oral Bacteria." Microbiology and Molecular Biology Reviews, vol. 66, no. 3, 2002, pp. 486–505.
Loe, H. "Oral Hygiene in the Prevention of Caries and Periodontal Disease." International Dental Journal, vol. 50, no. 3, 2000, pp. 129–139.
Oyanagi, T., et al. "Potentials of Mouthwashes in Disinfecting Cariogenic Bacteria and Biofilms Leading to Inhibition of Caries." The Open Dentistry Journal, vol. 6, 2012, pp. 23–30.
Crevelin, E.J., et al. "Antimicrobial Activity of the Essential Oil of Plectranthus neochilus Against Cariogenic Bacteria." Evidence-Based Complementary and Alternative Medicine, vol. 2015, 2015, p. 102317.
Wu, Y.T., et al. "Pharmacokinetic Studies of Chinese Medicinal Herbs Using an Automated Blood Sampling System and Liquid Chromatography-Mass Spectrometry." Journal of Traditional and Complementary Medicine, vol. 2, no. 1, 2012, pp. 33–40.
Vasconcelos, N.G., et al. "Antibacterial Mechanisms of Cinnamon and Its Constituents: A Review." Microbial Pathogenesis, vol. 120, 2018, pp. 198–203.
Adams, T.B., et al. "The FEMA GRAS Assessment of Cinnamyl Derivatives Used as Flavor Ingredients." Food and Chemical Toxicology, vol. 42, 2004, pp. 157–185. https://doi.org/10.1016/j.fct.2003.08.021.
Idbeis, Hasan I. "Phenotypic Characterization and Phylogenetic Analysis of Streptococcus Mutans Strains Isolated from Dental Caries Patients." IAR Journal of Parasitology and Infectious Diseases, vol. 2, no. 1, 2023, pp. 1–6.
Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Second Informational Supplement. CLSI Document M100-S22, Wayne, PA, 2012.
Kwasny, S. and T. Opperman. "Static Biofilm Cultures of Gram-Positive." Current Protocols in Pharmacology, 2010.
Belli, W.A. and R.E. Marquis. "Adaptation of Streptococcus Mutans and Enterococcus hirae to Acid Stress in Continuous Culture." Applied and Environmental Microbiology, vol. 57, 1991, pp. 1134–1138.
Sun, Y., et al. "Effects of Sub-Minimum Inhibitory Concentrations of Lemon Essential Oil on the Acid Tolerance and Biofilm Formation of Streptococcus Mutans." Archives of Oral Biology, vol. 87, 2018, pp. 235–241.
He, Z., et al. "Antimicrobial Activity of Cinnamaldehyde on Streptococcus Mutans Biofilms." Frontiers in Microbiology, vol. 10, 2019, p. 2241.
Jia, P., et al. "Effect of Cinnamaldehyde on Biofilm Formation and sarA Expression by Methicillin-Resistant Staphylococcus aureus." Letters in Applied Microbiology, vol. 53, no. 4, 2011, pp. 409–416.
Kouidhi, B., et al. "Drug Resistance of Bacterial Dental Biofilm and the Potential Use of Natural Compounds as Alternative for Prevention and Treatment." Microbial Pathogenesis, vol. 80, 2015, pp. 39–49. https://doi.org/10.1016/j.micpath.2015.02.007.
Polaquini, C.R., et al. "Antibacterial and Antitubercular Activities of Cinnamylideneacetophenones." Molecules, vol. 22, 2017, p. E1685. https://doi.org/10.3390/molecules22101685.
Ribeiro, M., et al. "Cytotoxicity and Antimicrobial Action of Selected Phytochemicals Against Planktonic and Sessile Streptococcus Mutans." PeerJ, vol. 6, 2018, p. e4872. https://doi.org/10.7717/peerj.4872.
Pandit, S., et al. "Effects of Withania somnifera on the Growth and Virulence Properties of Streptococcus Mutans and Streptococcus sobrinus at Sub-MIC Levels." Anaerobe, vol. 19, 2013, pp. 1–8. https://doi.org/10.1016/j.anaerobe.2012.10.007.
Gregoire, S., et al. "Influence of Cranberry Phenolics on Glucan Synthesis by Glucosyltransferases and Streptococcus Mutans Acidogenicity." Journal of Applied Microbiology, vol. 103, 2007, pp. 1960–1968. https://doi.org/10.1111/j.1365-2672.2007.03441.x.
Gabe, V., et al. "Inhibitory Effects of Ethyl Gallate on Streptococcus Mutans Biofilm Formation by Optical Profilometry and Gene Expression Analysis." Molecules, vol. 24, 2019, p. E529. https://doi.org/10.3390/molecules24030529.
De, A., "Deficiency of MecA in Streptococcus Mutans Causes Major Defects in Cell Envelope Biogenesis, Cell Division and Biofilm Formation." Frontiers in Microbiology, vol. 9, 2018, p. 2130. https://doi.org/10.3389/fmicb.2018.02130.
Veloz, J.J., et al. "Polyphenol-Rich Extract from Propolis Reduces the Expression and Activity of Streptococcus Mutans Glucosyltransferases at Subinhibitory Concentrations." Biomedical Research International, vol. 2016, 2016, p. e4302706. https://doi.org/10.1155/2016/4302706.
Yamashita, Y., et al. "Role of the Streptococcus Mutans gtf Genes in Caries Induction in the Specific-Pathogen-Free Rat Model." Infection and Immunity, vol. 61, 1993, pp. 3811–3817.
Duque, C., et al. "Downregulation of GbpB, a Component of the VicRK Regulon, Affects Biofilm Formation and Cell Surface Characteristics of Streptococcus Mutans." Infection and Immunity, vol. 79, 2011, pp. 786–796. https://doi.org/10.1128/IAI.00725-10.
Fujita, K., et al. "Correlation of Biological Properties with Glucan-Binding Protein B Expression Profile in Streptococcus Mutans Clinical Isolates." Archives of Oral Biology, vol. 56, 2011, pp. 258–263. https://doi.org/10.1016/j.archoralbio.2010.09.018.
Burgess, N.A., et al. "LuxS-Dependent Quorum Sensing in Porphyromonas gingivalis Modulates Protease and Haemagglutinin Activities but Is Not Essential for Virulence." Microbiology, vol. 148, no. 3, 2002, pp. 763–772.
Gutierrez, J.A., et al. "Streptococcus Mutans ffh, a Gene Encoding a Homologue of the 54 kDa Subunit of the Signal Recognition Particle, Is Involved in Resistance to Acid Stress." Microbiology, vol. 145, no. 2, 1999, pp. 357–366.
Leung, V., et al. "Death and Survival in Streptococcus Mutans: Differing Outcomes of a Quorum-Sensing Signaling Peptide." Frontiers in Microbiology, vol. 6, 2015, p. 1176.
License
Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License
All papers should be submitted electronically. All submitted manuscripts must be original work that is not under submission at another journal or under consideration for publication in another form, such as a monograph or chapter of a book. Authors of submitted papers are obligated not to submit their paper for publication elsewhere until an editorial decision is rendered on their submission. Further, authors of accepted papers are prohibited from publishing the results in other publications that appear before the paper is published in the Journal unless they receive approval for doing so from the Editor-In-Chief.
Himalayan Journal of Applied Medical Sciences and Research open access articles are licensed under a Creative Commons Attribution-Share A like 4.0 International License. This license lets the audience to give appropriate credit, provide a link to the license, and indicate if changes were made and if they remix, transform, or build upon the material, they must distribute contributions under the same license as the original.
Advertisement
Recommended Articles
Research Article
Association Between Oxidative Stress Biomarkers and Age-Related Hearing Loss: A Hospital-Based Case–Control Study
Watheq A. Mohammed Ali,
...
Mohammed Qasim Mohammed
Published: 24/01/2026
Download PDF
Cite
x
APA
Mohammed Ali, W. A., Hussien Dabi, A. & Qasim Mohammed, M. (2026). Association Between Oxidative Stress Biomarkers and Age-Related Hearing Loss: A Hospital-Based Case–Control Study. Himalayan Journal of Applied Medical Sciences and Research, 7(1), 1-5.
MLA
Mohammed Ali, Watheq A., Abdulkareem Hussien Dabi and Mohammed Qasim Mohammed. "Association Between Oxidative Stress Biomarkers and Age-Related Hearing Loss: A Hospital-Based Case–Control Study." Himalayan Journal of Applied Medical Sciences and Research 7.1 (2026): 1-5.
Chicago
Mohammed Ali, Watheq A., Abdulkareem Hussien Dabi and Mohammed Qasim Mohammed. "Association Between Oxidative Stress Biomarkers and Age-Related Hearing Loss: A Hospital-Based Case–Control Study." Himalayan Journal of Applied Medical Sciences and Research 7, no. 1 (2026): 1-5.
Harvard
Mohammed Ali, W. A., Hussien Dabi, A. and Qasim Mohammed, M. (2026) 'Association Between Oxidative Stress Biomarkers and Age-Related Hearing Loss: A Hospital-Based Case–Control Study' Himalayan Journal of Applied Medical Sciences and Research 7(1), pp. 1-5.
Vancouver
Mohammed Ali WA, Hussien Dabi A, Qasim Mohammed M. Association Between Oxidative Stress Biomarkers and Age-Related Hearing Loss: A Hospital-Based Case–Control Study. Himalayan Journal of Applied Medical Sciences and Research. 2026 Jan;7(1):1-5.
Download PDF
Research Article
Bones in Balance: Awareness of Rickets and Nutritional Deficiencies Among the Hamirpur Community
Vishal Dhatwalia,
Swati Chandel
Published: 10/12/2024
Download PDF
Cite
x
APA
Dhatwalia, V. & Chandel, S. (2024). Bones in Balance: Awareness of Rickets and Nutritional Deficiencies Among the Hamirpur Community. Himalayan Journal of Applied Medical Sciences and Research, 5(2), 1-5.
MLA
Dhatwalia, Vishal and Swati Chandel. "Bones in Balance: Awareness of Rickets and Nutritional Deficiencies Among the Hamirpur Community." Himalayan Journal of Applied Medical Sciences and Research 5.2 (2024): 1-5.
Chicago
Dhatwalia, Vishal and Swati Chandel. "Bones in Balance: Awareness of Rickets and Nutritional Deficiencies Among the Hamirpur Community." Himalayan Journal of Applied Medical Sciences and Research 5, no. 2 (2024): 1-5.
Harvard
Dhatwalia, V. and Chandel, S. (2024) 'Bones in Balance: Awareness of Rickets and Nutritional Deficiencies Among the Hamirpur Community' Himalayan Journal of Applied Medical Sciences and Research 5(2), pp. 1-5.
Vancouver
Dhatwalia V, Chandel S. Bones in Balance: Awareness of Rickets and Nutritional Deficiencies Among the Hamirpur Community. Himalayan Journal of Applied Medical Sciences and Research. 2024 Jul;5(2):1-5.
Download PDF
Research Article
Clinical and Functional Outcomes of Interlocking and Küntscher Nailing in the Management of Femoral Shaft Fractures: A Prospective Study
Ahmed Abdalzahra Mohaisen ,
...
Mohammed Saad Abdulzahra
Published: 10/01/2026
Download PDF
Cite
x
APA
None, A. A. M., None, H. A. A. & None, M. S. A. (2026). Clinical and Functional Outcomes of Interlocking and Küntscher Nailing in the Management of Femoral Shaft Fractures: A Prospective Study. Himalayan Journal of Applied Medical Sciences and Research, 7(1), 1-7.
MLA
None, Ahmed Abdalzahra Mohaisen, Hassan Ali Abid and Mohammed Saad Abdulzahra . "Clinical and Functional Outcomes of Interlocking and Küntscher Nailing in the Management of Femoral Shaft Fractures: A Prospective Study." Himalayan Journal of Applied Medical Sciences and Research 7.1 (2026): 1-7.
Chicago
None, Ahmed Abdalzahra Mohaisen, Hassan Ali Abid and Mohammed Saad Abdulzahra . "Clinical and Functional Outcomes of Interlocking and Küntscher Nailing in the Management of Femoral Shaft Fractures: A Prospective Study." Himalayan Journal of Applied Medical Sciences and Research 7, no. 1 (2026): 1-7.
Harvard
None, A. A. M., None, H. A. A. and None, M. S. A. (2026) 'Clinical and Functional Outcomes of Interlocking and Küntscher Nailing in the Management of Femoral Shaft Fractures: A Prospective Study' Himalayan Journal of Applied Medical Sciences and Research 7(1), pp. 1-7.
Vancouver
Ahmed Abdalzahra Mohaisen AAM, Hassan Ali Abid HAA, Mohammed Saad Abdulzahra MSA. Clinical and Functional Outcomes of Interlocking and Küntscher Nailing in the Management of Femoral Shaft Fractures: A Prospective Study. Himalayan Journal of Applied Medical Sciences and Research. 2026 Jan;7(1):1-7.
Download PDF
Research Article
A Critical Analysis of Spinal Block Anesthesia under Ultrasonographic Guided Technique: Is Levobupivacaine Alone Effective or on Adjuvant is Warranted?
Verma AK
Published: 20/09/2021
Download PDF
Cite
x
APA
AK, V. (2021). A Critical Analysis of Spinal Block Anesthesia under Ultrasonographic Guided Technique: Is Levobupivacaine Alone Effective or on Adjuvant is Warranted?. Himalayan Journal of Applied Medical Sciences and Research, 2(2), 1-4.
MLA
AK, Verma. "A Critical Analysis of Spinal Block Anesthesia under Ultrasonographic Guided Technique: Is Levobupivacaine Alone Effective or on Adjuvant is Warranted?." Himalayan Journal of Applied Medical Sciences and Research 2.2 (2021): 1-4.
Chicago
AK, Verma. "A Critical Analysis of Spinal Block Anesthesia under Ultrasonographic Guided Technique: Is Levobupivacaine Alone Effective or on Adjuvant is Warranted?." Himalayan Journal of Applied Medical Sciences and Research 2, no. 2 (2021): 1-4.
Harvard
AK, V. (2021) 'A Critical Analysis of Spinal Block Anesthesia under Ultrasonographic Guided Technique: Is Levobupivacaine Alone Effective or on Adjuvant is Warranted?' Himalayan Journal of Applied Medical Sciences and Research 2(2), pp. 1-4.
Vancouver
AK V. A Critical Analysis of Spinal Block Anesthesia under Ultrasonographic Guided Technique: Is Levobupivacaine Alone Effective or on Adjuvant is Warranted?. Himalayan Journal of Applied Medical Sciences and Research. 2021 Jul;2(2):1-4.
Ikareim Idbeis, H., R. Arian AL-Aidy, S., Abbas Gibar Aifar, S. & Bahlok Jebur, H. (2023). Inhibitory Effects of Cinnamaldehyde on Biofilm Formation and Virulence Factors in Streptococcus Mutans. Himalayan Journal of Applied Medical Sciences and Research, 4(2), 1-8.
MLA
Ikareim Idbeis, Hasan, et al. "Inhibitory Effects of Cinnamaldehyde on Biofilm Formation and Virulence Factors in Streptococcus Mutans." Himalayan Journal of Applied Medical Sciences and Research 4.2 (2023): 1-8.
Chicago
Ikareim Idbeis, Hasan, Salem R. Arian AL-Aidy, Sagia Abbas Gibar Aifar and Hakim Bahlok Jebur. "Inhibitory Effects of Cinnamaldehyde on Biofilm Formation and Virulence Factors in Streptococcus Mutans." Himalayan Journal of Applied Medical Sciences and Research 4, no. 2 (2023): 1-8.
Harvard
Ikareim Idbeis, H., R. Arian AL-Aidy, S., Abbas Gibar Aifar, S. and Bahlok Jebur, H. (2023) 'Inhibitory Effects of Cinnamaldehyde on Biofilm Formation and Virulence Factors in Streptococcus Mutans' Himalayan Journal of Applied Medical Sciences and Research 4(2), pp. 1-8.
Vancouver
Ikareim Idbeis H, R. Arian AL-Aidy S, Abbas Gibar Aifar S, Bahlok Jebur H. Inhibitory Effects of Cinnamaldehyde on Biofilm Formation and Virulence Factors in Streptococcus Mutans. Himalayan Journal of Applied Medical Sciences and Research. 2023 Jul;4(2):1-8.